
MONDAY, AUGUST 23, 2021. BY STAN GRANT, VITICULTURIST.
For many wine drinkers, the most important characteristic of a wine is its flavor. While sugar (sweetness and alcohol strength), acidity (tartness), and color related compounds (astringency and bitterness) in grapes contribute to wine flavor, the taste characteristics most unique to a specific wine are usually associated with distinct aromas. We humans can sense and distinguish minute amounts of the volatile compounds contributing to the bouquet of a wine.

Figure 1. Many aroma, flavor, and color compounds in wines originate in grapes.
Of course, for vineyard managers, the focus is the aroma, flavor, color, and other sensory compounds in the berries in their vineyards prior to delivery to the winery (Figure 1). In recent decades, extensive research has deepened our understanding of sensory compounds, including their development in fruit and how environmental factors influence them. In spite of the progress, managing vineyards to the best advantage of sensory compounds can be challenging for several reasons, including the following.
During the growing season, the array of sensory compounds in berries continually changes as various molecules are synthesized or degraded. In other words, fruit composition is a moving target. For some sensory compounds, seasonal changes are somewhat independent of fruit maturation and difficult to predict. Sensory compounds are hard to monitor in berries because they occur at very low concentrations or they occur in bound forms not sensible until freed during processing at the winery. Finally, prevailing growing season weather may profoundly impact sensory compound concentrations; sometimes obscuring the effects of practices we have implemented to enhance them. In fact, nearly anything that directly or indirectly affects grapevine metabolism and growth can influence sensory compounds and their intensity in fruit, with vintage growing conditions being paramount.
Here we will focus upon three groups of sensory compounds that we may impact in vineyards. They are monoterpenes, methoxypyrazines, and phenolics. Traditionally, plant scientists have referred to these and other organic molecules perceived to be non-essential for plant growth and development as secondary metabolites. Secondary metabolites, including those listed above, vary in type and concentration among grapevine species and varieties.
Floral Characters – Monoterpenes
The fruit of all wine grape varieties contain volatile terpene (isoprenoid) compounds, but they are usually minor contributors to varietal character. There are, however, a few varieties in which a specific group of terpenes are major players. Monoterpenes, the first aroma compounds characterized in grape berries, impart floral qualities to the fruit of Gewurztraminer, the Muscat family of varieties, and the Musque clones of Chardonnay and Sauvignon blanc. They are also present in lesser amounts in Riesling, Chenin blanc, and Viognier berries. Rose, lilac, pine, peach, and citrus are among the descriptors for the floral qualities of monoterpenes.
Two pools of monoterpenes occur in berries. They are the free volatile terpenes (FVT), which are readily sensible compounds, and the potential volatile terpenes (PVT), which are insensible, bound compounds. In berries, the content of bound monoterpenes, many of which become sensible during winemaking, is greater than that of free monoterpenes. Like most aroma molecules, monoterpenes are most concentrated in berry skins, but they also occur at lower concentrations in the pulp and juice. Monoterpenes first appear in berries before veraison and they normally increase late into ripening.

Figure 2. Severe water stress causes a loss of volatile aroma compounds from berries. (Photo Source: Progressive Viticulture©)
Cool climate research involving fruit zone leaf removal has demonstrated the importance of sunlight exposure on monoterpene synthesis. In warm climates, however, direct sunlight exposure can diminish monoterpene concentrations, perhaps due to berry heating. Under these conditions, a thin veneer of leaves, consisting of a single leaf layer, on the afternoon sun side of fruit zones favor maximum monoterpene concentration in berries. To further this effort, manage vineyard water to avoid severe stress, which is associated with a loss of potential aroma (Figure 2).
Green Aromas – Methoxypyrazines
Among the best understood sensory compounds are the methoxypyrazines, which contribute to vegetative aromas to berries and wine. There are two principal methoxypyrazine compounds. One methoxypyrazine compound (IBMP) is associated with bell pepper, green bean, and green gooseberry characters, while the other (IPMP) is associated with peas and cooked asparagus. Excessive vegetative characters associated with these compounds are considered unpleasant and a mark of underripe fruit.
Methoxypyrazines are prevalent in certain varieties, including Cabernet Sauvignon, Cabernet Franc, Merlot, Sauvignon blanc, and Semillon, but absent in others. They occur in berry skins and seeds, as well as in stems and leaves. Methoxypyrazines are particularly abundant in mature leaves.
Methoxypyrazines reach their greatest concentrations in berries prior to veraison and their concentrations decline thereafter. Their rate of decline depends on growing conditions. Cool growing seasons and cluster shading slow methoxypyrazine degradation, often allowing pronounced vegetative characters to persist in otherwise ripe fruit.

Figure 3. Reducing canopy density with fruit zone leaf and lateral shoot removal prior to berry touch also reduces vegetative characteristics in berries. (Photo Source: Progressive Viticulture©)
Grape growers can enhance methoxypyrazine loss by implementing practices that promote fruit exposure to dappled sunlight and a favorable crop load (leaf area per unit fruit). These include leaf removal and/or lateral shoot removal from fruit zones between fruit set and the beginning of berry touch (Eichhorn-Lorenz stages 27 and 33, respectively), and if needed, in-season hedging of the apical ends of shoots to ensure no more than about 14 to 20 well-exposed, healthy leaves per shoot bearing 2 clusters (Figure 3).
The efficacy of both leaf removal and hedging benefit from dormant season pruning and prebloom shoot thinning leading to about 5 to 6 shoots per foot cordon or cane, as well as regulated deficit irrigation schedules that maintain healthy foliage while preventing renewed shoot growth. In addition, it is important to harvest grapes when fully ripe and to minimize stems and leaves among harvested grapes.
Color, Astringency, Bitterness, and Aroma – Phenolics
Phenolics are a highly diverse and complex group of compounds having an assortment of sensory affects. Although they are present at comparatively small concentrations, phenolics are the third most plentiful organic compounds in berries (after carbohydrates and acids).
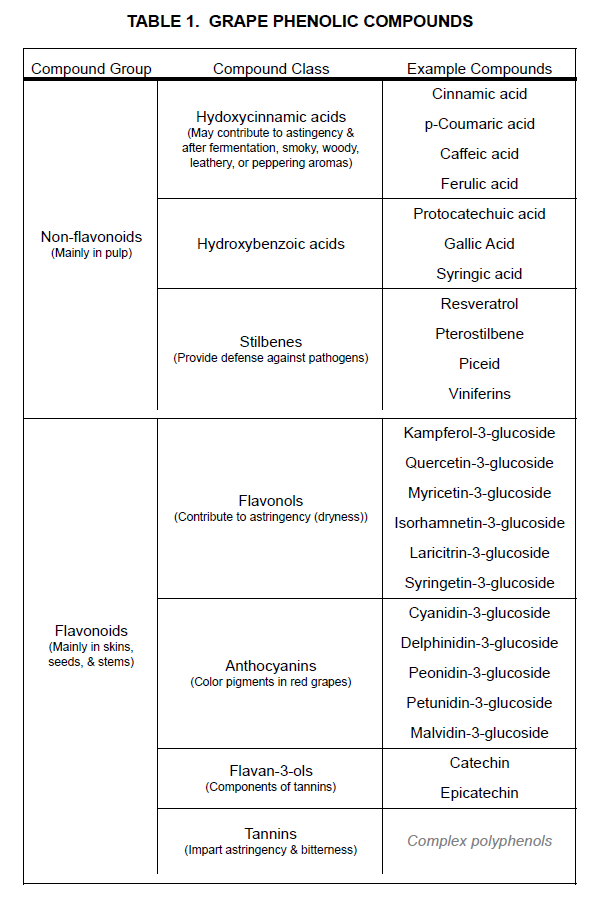
There are two primary groups of phenolic compounds in grapes. They are the flavonoids, which accumulate mainly in berry skins, seeds, and stems, and the non-flavonoids, which accumulate mainly in the pulp of berries (Table 1). Among flavonoids are two that are particularly important for red grapes: tannins that impart astringency (dryness) and bitterness, and anthocyanins that impart red, blue, and purple color. For white varieties, the inability to produce anthocyanins has only a small impact on the concentrations of other phenolic compounds. Aromas, such as vanilla, smoke, wood, leather, and pepper, are among those associated with phenolic compounds.
In berries, phenolics decrease until veraison. For white varieties, the general phenolic decline continues during ripening, although specific volatile phenolic compounds may increase, stay the same, or decrease. In contrast, red berry phenolics steadily increase after veraison largely due to anthocyanin production.

Figure 4. High temperatures inhibit fruit coloration and in severe cases, leave fruit with a bleached appearance. (Photo Source: Progressive Viticulture©)
Sunlight or at least, limited shade promotes the synthesis and accumulation of most phenolics, including anthocyanins and flavonols. However, high temperatures, which are often associated with excessive sunlight, diminish phenolic concentrations (Figure 4). This reduction is due to both decreased phenolic synthesis and oxidation of existing phenolics.
In addition to the growing environment, the total phenolic content in berries depends on the variety. While there are variations in total phenolics among red varieties and clones, the difference between red and white varieties is more marked. This difference is the result of the preponderance of phenolic compounds in the skins of red grapes. Phenolic differences between red and white become more striking for wines due to dissimilarity in their processing. Phenolics in skins, seeds, and stems are extracted during red winemaking, while removal of skins, seeds, and stems from white musts prior to fermentation precludes phenolic extraction from them.
It is a happy coincidence that vineyard attributes that enhance phenolics in berries are the same as those listed above for limiting methoxypyrazines. They include fruit exposure to dappled sunlight, the balance between fruit and exposed healthy leaves, and moderate grapevine water stress all achieved as soon as possible after the fruit is set and maintained through ripening.
At the same time, ensure an adequate and balanced supply of mineral nutrients to support normal leaf and root activity, inhibit premature leaf senescence, and promote phenolic production. Deficient boron, a phenolic synthesis cofactor, diminishes phenolic intensity including the color of red grapes. Phosphorus deficiency also decreases anthocyanins. Both deficient and excess grapevine nitrogen decrease fruit phenolics. Ample nitrogen, on the other hand, may moderate bitterness and astringency in white grapes.

Figure 5. Anthocyanins, the phenolic compounds responsible for color in red grapes, decline as fruit overripens. (Photo Source: Progressive Viticulture©)
Picking of ripe fruit with negligible leaves and stems is also part of vineyard management for a desirable phenolic profile (Figure 5). Following harvest, deliver fruit to the winery as soon as possible and, if feasible, transport it under cool conditions. Doing so will minimize the oxidation of phenolic compounds leading to browning, which is especially a concern for white berries.
In Summary
Called secondary compounds because they appear non-essential for grapevine survival, sensory metabolites are minor constituents of grapes, but major players for wine drinkers. Optimizing their content in berries involves three in-season vineyard attributes achieved soon after fruit set – dappled sunlight in fruit zones, fruit and functional leaf area balance, and moderate vine water stress. Equally important, the best possible fruit quality requires proper harvest timing to capture desirable sensory characteristics. During harvest, minimize leaves and stems from grape loads, and promptly deliver fruit to the winery in the cool part of the day.
A version of this article was originally published in the Mid Valley Agricultural Services September 2006 newsletter and was updated for this blog post.
Further Reading
Allen, MM. Contribution of methoxypyrazines to the flavor Cabernet Sauvignon and Sauvignon blanc. In PJ Williams (ed.) Proceedings of the Seventh annual Australian Wine Industry Technical Conference, Adelaide. 1990.
Augustyn, PH, Rapp, A, and Van Wyk, CJ. Some volatile aroma components of Vilis vinifera L. ev. Sauvignon blanc. South African Journal of Enology and Viticulture. 3, 53-60. 1982.
Belancic, A, Agosin, E, Ibacache, A, Bordeu, E, Baumes, R, Razungles, A, and Bayonove, C. Influence of sun exposure on the aromatic composition of Chilean muscat grape cultivars Moscatel del Alejandria and Moscatel rosada. American Journal of Enology & Viticulture. 48, 181-186. 1997.
Bisson, L. In search of optimal grape maturity. Practical Winery & Vineyard. 23,32,34,36,38,40,42-43. July/August 2001.
Calderon-Orellana, A, Matthews, MA, Drayton, WM, and Shackel, KA. Uniformity of ripeness and size in Cabernet Sauvignon berries from vineyards with contrasting crop price. American Journal of Enology & Viticulture. 65: 81-88. 2014.
Chapman, DM, Matthews, MA, and Guinard J-X. Sensory attributes of Cabernet Sauvignon wines made from vines with different crop yields. American Journal of Enology & Viticulture. 55: 325-334. 2004.
Downey, MO, Dokoozlian, NK, and Kristic, MP. Cultural practice and environmental impacts on flavonoid composition of grapes and wine: A review of recent research. American Journal of Enology & Viticulture. 57, 257-268. 2006.
Dunlevy, JD, Soole, KL, Perkins, MV, Nicholson, EL, Maffei, SM, and Boss, PK. Determining the methoxypyrazine biosynthesis variables affected by light exposure and crop level in Cabernet Sauvignon. American Journal of Enology & Viticulture. 64: 450-458. 2013.
Galet, P. General Viticulture. Oneoplurimedia, Chateau de Chaintre, France, 2000.
Grant, S. Extended wine grape ripening. Practical Winery and Vineyard. 38 (3); 26-37. Jul/Aug 2006.
Hamilton, RP, and Coombe, BG. Harvesting of winegrapes. In Coombe, BG and Dry, PR (ed.). Viticulture. Volume 2 Practices. Winetitles, Adelaide, 1992.
Hashizume, K, and Samuta, T. Grape maturity and light exposure affect berry methoxypyrazine concentration. American Journal of Enology & Viticulture. 50: 194-198. 1999.
Jackson, DI, and Lombard, PB. Environmental and management practices affecting grape composition and wine quality – A review. American Journal of Enology & Viticulture. 44, 409-430. 1993.
Keller, M, Pool, RM, and Henick-Kling, T. Excessive nitrogen supply and shoot trimming can impair color development in Pinot noir grapes and wine. Australian Journal of Grape and Wine Research. 5, 45-55. 1999.
Keller, M. The science of grapevines. Academic Press, Burlington, MA. 2010.
Kennedy, JA, Matthews, MA, and Waterhouse, AL. Effect of maturity and vine water status on grape skin and wine flavonoids. American Journal of Enology & Viticulture. 53, 268-274. 2002.
Kennedy, JA, Saucier, C, and Glories, Y. Grape, and wine phenolics; history and perspective. American Journal of Enology & Viticulture. 57, 239-248. 2006.
Koch, A, Doyle, CL, Matthews, MA, Williams, LE, and Ebeler, SE. 2-methoxy-3-isobutylpyrazine in grape berries and its dependence on genotype. Phytochemistry. 71, 2190-2198. 2010.
Koch, A, Ebeler, SE, Williams, LE, and Matthews, MA. Fruit ripening in Vitis vinifera: light intensity before and not during ripening determines the concentration of 2-methoxy-3-isobutylpyrazine in Cabernet Sauvignon. Physiologia Plantarum. 145, 275-285. 2012.
Lacey, MJ, Allen, MS, Harris, RLN, and W. Brown, WV. Methoxypyrazines in Sauvignon blanc grapes and wines. American Journal of Enology & Viticulture. 42, 103-108. 1991.
Marais, J, Hunter, JJ, and Haasbroek, PD. Effect of canopy microclimate, season, and region on Sauvignon blanc grape composition and wine quality. South African Journal of Enology and Viticulture. 20, 19-30. 1999.
Matthews, MA, Ishii, R, Anderson, MM, and O’Mahony, M. Dependence of wine sensory attributes on vine water status. Journal of the Science of Food & Agriculture. 51: 321-335. 1990.
Matthews, M. Water deficits, yield, and berry size as factors of composition and wine sensory attributes. In Williams, L. (ed.). Proc. 7th Int. Symp. Grapevine Physiol. Biotech. p. 94. Davis, Calif. June 2004.
Mullins, MG, Bouquet, A, and Williams, LE. Biology of the Grapevine. Cambridge University Press, Cambridge. 1992.
Reynolds, AG, and Wardle, DA. Flavour development in the vineyard: impact of viticultural practices on grape monoterpenes and their relationship to wine sensory response. South African Journal of Enology and Viticulture. 18, 3-18. 1997.
Robinson, AL, Boss, PK, Solomon, PS, Trengove, RD, Heymann, H, and Ebler, SE. Origins of grape and wine aroma. Part 1. Chemical components and viticultural impacts. American Journal of Enology & Viticulture. 65: 1-24. 2014.
Rotter, B. Fruit maturity assessment. Improved winemaking (members.tripod.com/~BRotter/)
Roujou de Boube ́e, D, Van Leeuwen, C, and Dubourdieu, D. Organoleptic impact of 2-methoxy-3- isobutylpyrazine on red Bordeaux and Loire wines. Effect of environmental conditions on concentrations in grapes during ripening. Journal of Agriculture & Food Chemistry. 48: 4830–4834. 2000.
Skinner, PW, Ishii, R, O’Mahony, M, and Matthews, MA. Implications of fertilizer additions to vines growing under phosphorus-deficient conditions on different soil types with regard to wine sensory attributes. Oeno One. 2, 129-146. 2019.
Ward, SC, Petrie, PR, Johnson, TE, Boss, PK, and Bastian, SEP. Unripe berries and petioles in Vitis vinifera cv. Cabernet Sauvignon fermentations affect sensory and chemical profiles. American Journal of Enology & Viticulture. 66: 435-443. 2015.
Winter, E. Avoiding “green” characters in Cabernet Sauvignon. Practical Winery and Vineyard. May/June 2004.
Have something interesting to say? Consider writing a guest blog article!
To subscribe to the Coffee Shop Blog, send an email to stephanie@lodiwine.com with the subject “blog subscribe.”
To join the Lodi Growers email list, send an email to stephanie@lodiwine.com with the subject “grower email subscribe.”
To receive Lodi Grower news and event promotions by mail, send your contact information to stephanie@lodiwine.com or call 209.367.4727.
For more information on the wines of Lodi, visit the Lodi Winegrape Commission’s consumer website, lodiwine.com.
For more information on the LODI RULES Sustainable Winegrowing Program, visit lodigrowers.com/standards or lodirules.org.
