 Many species of microorganisms may infect grape berries, including fungi and bacteria. Several species of these fungi, including Botrytis cinerea, Aspergillus niger, A. carbonarius, Cladosporium cladosporioides, C. herbarum, Penicillium spp., Rhizopus arrizhus, and Alternaria spp., participate in bunch rot of grapes, though the latter four species are not true pathogens but rather secondary colonizers. Botrytis cinerea may directly penetrate and infect healthy berries, but the others require wounds or previously infected berries to infect the fruit. Economic losses result from the direct reduction in yield as the rotted fruit is culled and from the increased harvest costs associated with this practice. Table and raisin grape quality and storage potential, as well as wine quality, are all negatively affected by bunch rots. Losses can be extremely high in some years, especially in locations where conditions for B. cinerea are extremely favorable. Botrytis bunch rot is the principal fruit rot disease of grapes grown in the production areas of the coast and the northern San Joaquin Valley subject to the marine influence of the San Francisco Bay. In the warmer central and southern San Joaquin Valley, summer bunch rot or sour rot is the more common fruit disease. However, in coastal areas during warm summers it is not uncommon to see sour rot occurring, particularly on Zinfandel but on other cultivars as well.
Many species of microorganisms may infect grape berries, including fungi and bacteria. Several species of these fungi, including Botrytis cinerea, Aspergillus niger, A. carbonarius, Cladosporium cladosporioides, C. herbarum, Penicillium spp., Rhizopus arrizhus, and Alternaria spp., participate in bunch rot of grapes, though the latter four species are not true pathogens but rather secondary colonizers. Botrytis cinerea may directly penetrate and infect healthy berries, but the others require wounds or previously infected berries to infect the fruit. Economic losses result from the direct reduction in yield as the rotted fruit is culled and from the increased harvest costs associated with this practice. Table and raisin grape quality and storage potential, as well as wine quality, are all negatively affected by bunch rots. Losses can be extremely high in some years, especially in locations where conditions for B. cinerea are extremely favorable. Botrytis bunch rot is the principal fruit rot disease of grapes grown in the production areas of the coast and the northern San Joaquin Valley subject to the marine influence of the San Francisco Bay. In the warmer central and southern San Joaquin Valley, summer bunch rot or sour rot is the more common fruit disease. However, in coastal areas during warm summers it is not uncommon to see sour rot occurring, particularly on Zinfandel but on other cultivars as well.
__________________________________________________________________________________
This excerpt from Grape Pest Management (Third Edition) was published with permission from UC ANR, and was written by Larry Bettiga and Douglas Gubler.
The Lodi Winegrape Commission and the University of California Division of Agriculture and Natural Resources (UC ANR) are partnering to provide Lodi winegrowers with the latest information about grape pest management. This is the first of several excerpts from the third edition of the Grape Pest Management book to be published here in the Coffee Shop.

In Grape Pest Management, more than 70 research scientists, cooperative extension advisors and specialists, growers, and pest control advisors have consolidated the latest scientific studies and research into one handy reference. The result is a comprehensive, easy-to-read pest management tool.
The new edition, the first to be published in over a decade, includes several new invasive species that are now major grape pests. It also reflects an improved understanding among researchers and growers about the biology of pests. With nine expansive chapters, helpful, colorful photos throughout, here’s more of what you’ll find:
-Diagnostic techniques for identifying vineyard problems
-Detailed descriptions of more than a dozen diseases
-Comprehensive, illustrated listings of insect and mite pests
-Regional calendars of events for viticultural management
-Up-to-date strategies for vegetation management
__________________________________________________________________________________

Figure 13.1 Rotting of young tissue characteristic of Botrytis shoot blight. Photo: J. K. Clark.
The most important fungal plant pathogen responsible for bunch rot of grape berries is Botrytis cinerea (Persoon). This fungus has a wide host range of both native and cultivated plants. On grape it can grow on any plant tissue that is succulent (young leaves and shoots or flower parts), stressed (damaged fruit), ripened fruit, or dead tissue (yellow leaves or senescent flower parts). The enzymes produced by the fungus can destroy the integrity of a grape berry within a few days. Botrytis bunch rot can severely affect tight-clustered, thin-skinned cultivars, especially under heavy canopies or during wet growing seasons. In the production of late-harvest wines, a late-season rot caused by B. cinerea is referred to as Noble rot and is highly desirable to produce a concentration of sugars and flavor components under the right environmental conditions for fruit processed into dessert wines.
Foliage Symptoms
In the spring, Botrytis can infect succulent shoots, flowers, or young leaves when prolonged warm, moist conditions caused by frequent rains occur. Patches of soft brown tissue develop, resulting in the death of the infected plant part. Small shoots can be killed by this type of infection, and it can reduce yields due to the loss of flower clusters. Older shoot stems may be girdled at the site of infection and wilt or break off (fig. 13.1). Infection of young expanded leaves can produce areas of brown necrotic tissue (fig. 13.2). In almost all cases, infection occurs in the axils of leaf or inflorescence junctions with the main shoot (figs. 13.3–13.4). The infection results from water pooling in these crotches and remaining for a long duration.

Figure 13.2 Spring Botrytis infection on leaf. Photo: L. J. Bettiga.
Fruit: Bunch rot often begins when blossoms become infected during rainfall. The pathogen invades the flower parts and becomes dormant until veraison. At veraison, individually infected berries in the cluster turn brown on white cultivars or reddish on red and black cultivars due to enzymes produced by the fungus and subsequent fungal invasion of the pulp (fig. 13.5). This stage is known as slip skin because fungal enzymes break down the cutin in the epidermis and it easily slips off the berry (fig. 13.6). If temperatures are moderate, moisture is high, and wind speed is low, epidermal cracks will form in which fungal growth produces mycelium and spores (fig. 13.7), resulting in the characteristic gray, velvety appearance of infected berries (fig. 13.8). The fungus can then spread from berry to berry causing a nested appearance of infected berries. If conditions remain favorable, the disease can result in a high percentage of berries being rotted (fig. 13.9), and if disease is severe enough to reach the rachis, raisining of infected berries can occur (fig. 13.10).
Canes: Poorly matured canes can become infected late in the growing season and show a bleaching of the bark. This whitening and the development of sclerotia can be seen during dormancy (fig. 13.11). Sclerotia are hard, black dormant structures about 3 mm (0.13 in) in diameter.

Figure 13.3 Botrytis lesion at node of young shoot in spring. Photo: J. Hall.
Disease Cycle
Overwintering: Botrytis survives winter by forming sclerotia either on the surface of or inside colonized plant tissue, including canes, rachises, and berries (fig. 13.12). The two most common sources of sclerotia in vineyards are grape mummy clusters from the previous season and canes that were infected in the fall of the year. After rains or irrigation in spring, the sclerotia germinate and produce masses of spores (conidia) (fig. 13.13), which are spread by air currents, workers, or rain splashing. The production of spores by B. cinerea results in the characteristic gray, fluffy appearance of the fungus on infected tissue (fig. 13.14).
Growing Season: To germinate and grow, spores of B. cinerea require continuous free water and nutrients for certain durations of time given certain temperatures. For example, at temperatures of 18° to 24°C (64.5° to 75°F), only 2 hours of free water are needed for germination to occur, while at temperatures lower than 16°C (61°F) and higher than 27°C (80.5°F), more time is needed for germination to occur. The free water can be from dew, fog, irrigation, rain, or juice from damaged berries. Periods as short as 15 minutes with no available water are sufficient to stop germination. However, even with water present, germination also requires nutrients, which can come from many sources, even from the surface of a healthy grape berry. The main nutrient required is a simple sugar, such as fructose or glucose. After veraison, Botrytis germination and infection can begin in a cluster where the berries have been damaged. Insects, birds, or mechanical damage can cause juice to be released, providing the spores with the nutrients and free water they need for germination. In coastal areas, this can result in a high incidence of Botrytis bunch rot.

Figure 13.4 Botrytis-blighted flower cluster. Photo: W. J. Moller.
Ambient air temperatures also affect germination. Warmer temperatures usually hasten the drying of the berries and directly reduce germination. Under field conditions, both temperature and duration of free moisture enable the fungus to infect. At and above 32°C (90°F), the fungus does not grow, but it will grow slowly even at 1°C (34°F), which allows it to continue to infect stored table grapes (see section 20, “Postharvest Diseases of Table Grapes”).
In early spring under moist conditions, the spores can infect the grape flowers, succulent young stems, or young leaves. During bloom, detached withering floral tissue (calyptras and stamens), aborted ovaries trapped within the developing cluster, and scar tissue on the receptacle (tip of the pedicel) left by the detachment of the calyptras can become infected and become a source of inoculum. These colonized tissues can give rise to spores when wet at any point later in the growing season.
Latent or quiescent berry infections can also occur when the style of the flower becomes infected during bloom. At veraison, as the sugar concentration increases the fungus resumes growth and spreads inside the berry. After veraison, sugar increases and the skins begin to soften; the berries themselves become more susceptible to infection. Rot can spread to adjoining berries in a cluster, or new infections can be initiated by airborne spores. Late-season infections are most severe when relative humidity exceeds 92%, free moisture is present on the fruit surface, and temperatures are in the 15° to 28°C (59° to 82.5°F) range. Research in controlled laboratory studies found a range in the proportion of berries infected when temperatures varied from 12° to 30°C (53.5° to 86°F) and the hours of leaf wetness varied from 4 to 20 hours. As little as 4 hours of surface moisture at optimal temperatures of 22°C (71.5°F) can lead to significant berry infection.

Figure 13.5 Infected berries turn a brown color on a white cultivar. Photo: L. J. Bettiga.
The fungus can penetrate grape berries through wounds; it can directly penetrate undamaged berries after veraison. The berry skin and epicuticular wax (known as bloom) are the main protection from infection by Botrytis. Any chemicals or cultural practices that alter these physical and chemical defenses will change the susceptibility of the berry to infection. For example, if fruit are grown in shady canopies with high relative humidity, the cuticle on the berries is generally significantly thinner and less wax is laid down, affording reduced protection from Botrytis infection. On the other hand, if fruit are exposed early, they develop a heavier cuticle and epicuticular wax layer and have more protection from infection.
In infected berries, cracks appear in the skin. The spores develop first in the cracks and then spread over the entire berry. The infection of other berries by the rapidly growing fungus and airborne spores contributes to the extremely rapid increase in disease observed after rains late in the season. Under optimal conditions, Botrytis can infect a berry, destroy it, and begin to produce spores in only 3 days. After infection, the berry may dry up if high temperatures and low relative humidities prevail. However, the fungus will still be alive and can continue to grow if favorable conditions for growth resume.
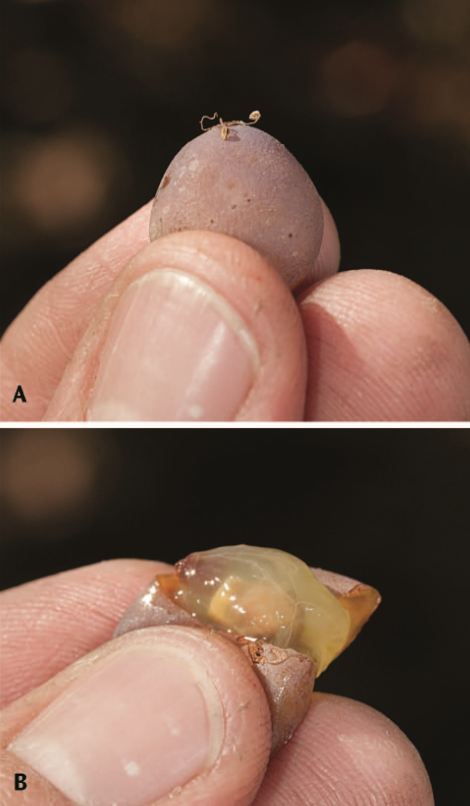
Figure 13.6 Breakdown of cutin in the epidermis (A) allows the pulp to be easily squeezed out (B). This is often referred to as the slip skin stage. Photos: L. J. Bettiga.
Feeding wounds caused by high populations of birds, the tortricid moths orange tortrix (Argyrotaenia franciscana) and omnivorous leafroller (Platynota stultana), or other berry-feeding insects can significantly increase the levels of bunch rot caused by Botrytis and other secondary fungal and bacterial pathogens. Berry scarring caused by early powdery mildew infections of developing berries can result in cracking during later growth stages, leading to increased levels of bunch rot. Warm, dry weather at harvest can desiccate Botrytis-infected berries (fig. 13.15).
Management
Control of Botrytis bunch rot is best achieved through an integrated approach that considers the management options listed below. The manipulation of the fruit zone to reduce humidity and promote cluster drying when preharvest rains occur is a cornerstone of the program. The efficacy of a fungicide, when needed, depends on getting good coverage, and coverage is affected by the canopy density and the stage of growth of the fruit clusters. By employing cultural control methods and properly applying fungicides as needed, the disease can be managed.
Sanitation: Sanitation is an important foundation for all effective disease control. Clusters left on vines or on the vineyard floor from the previous season can be a source of inoculum the following spring. Removing clusters from vines at pruning and placing them into the row middles where they can be incorporated into the soil can reduce inoculum.
Canopy Management: Since Botrytis spores have rigid environmental requirements for germination and growth, control can be obtained by creating a canopy microclimate that is less conducive to disease development. The objective is to expose the grape clusters to increased wind speed and light so that they dry out more quickly after a wetting.

Figure 13.7 Clusters of Botrytis spores on long stalks (conidiophores) on a berry surface. A Photo: L. J. Bettiga.
Canopy management can be directed to the canopy or to overall growth of the vine. Vineyard design decisions such as rootstock and scion selection, trellis type, training or pruning method, and plant spacing can affect canopy density. Row orientation can influence the fruit microclimatic parameters of wind speed, humidity, and temperature, which can alter the evaporative potential of the canopy and fruit. Careful planning to design a vineyard to the anticipated vigor of the site conditions will produce balanced canopies with moderate shoot vigor that optimizes leaf and cluster exposure that is appropriate for the site conditions. Vineyard design decisions generally have long-term effects on vine canopies that are not easily changed for the life of the vineyard. Other growing practices, such as irrigation and nutrition, can be altered annually; these practices can influence canopy density by increasing or decreasing the rate and amount of shoot and lateral growth. In general, the more dense the canopy (that is, the more layers of leaves surrounding the clusters), the more optimal conditions will be for development of Botrytis bunch rot. With careful management, it is often possible to reduce the density of the canopy, which in turn helps alleviate disease pressure.

Figure 13.8 Single infected berry and epidermis split on Riesling cluster (A); sporulation on infected berry (B). Photos: L. J. Bettiga.
Shoot thinning, shoot positioning, leaf removal, and hedging can increase air and light penetration to the clusters when canopies are too dense. Shoot thinning is a common practice for table and wine grape production. It can reduce density temporarily; the lateral shoot growth promoted by this practice can often increase canopy density by bloom to a level similar to a nonthinned vine. A light hedging is often used in vineyards in California to maintain an upright pattern of shoot growth that enhances air movement directly to the clusters. However, careful timing is needed to achieve satisfactory results. If hedging is used too early, lateral regrowth often makes the canopy denser than nonhedged vines, and in some cases these hedged vines have more bunch rot at harvest. Also, severe hedging removes a very large portion of the most photosynthetically active leaf area on the vine. This can result in delays of fruit maturity up to 3 weeks, offsetting any advantages of the open canopy for Botrytis control.
Removal of leaves and laterals around the clusters creates a microclimate within the canopy that is less conducive to development of Botrytis bunch rot. In many cases, the level of disease control obtained can equal or exceed the control from fungicide applications. When conducted immediately after fruit set, this practice can also physically shake off some of the floral debris that can be infected with Botrytis, reducing this source of inoculum. Leaf removal should be done on the side of the canopy that receives morning sun (east or north side) to reduce fruit damage from sunburn. In warmer areas, do not remove excessive numbers of leaves. If leaves are removed in the period after fruit set to when berries are BB-size, the berries acclimate readily to the sunlight and develop a thick cuticle and epicuticular wax that help prevent sunburn as well as Botrytis infection.

Figure 13.9 Destruction of the central part of a Chardonnay cluster caused by Botrytis (A). Desiccation and sporulation late in the growing season (B).Photos: L. J. Bettiga.
The microclimatic conditions in the canopy that most affect development of Botrytis are those that affect the duration of free water on the berries. The evaporation rate of water is affected by the temperature, light, vapor pressure deficit, and wind speed of the ambient air. When leaves around the clusters are removed, the wind speed around the clusters is increased, as are berry surface temperatures. These two factors contribute to drying the clusters after they have become wet.
In an unusually wet year, the benefits of microclimate modifications are reduced because of the impact of the macroclimate, and it may be necessary to increase applications of fungicides. If fungicides are required, however, proper canopy management will continue to be of benefit because more fungicide will be deposited on the more-exposed clusters.
Irrigation: Choosing the right type, timing, or level of irrigation can help control bunch rots. For example, overhead sprinkler irrigations near harvest can increase Botrytis levels. If this is the only type of irrigation available, vary the time of day or length of application to speed drying of the clusters. For example, in cool coastal valley vineyards, many growers irrigate at night to take advantage of the warmer, drier daytime conditions that aid drying. The length of time free moisture is on the clusters should not be greater than 15 hours, including the time it takes to dry the clusters completely.
Other types of irrigation should also be used judiciously. High levels of drip or furrow irrigation encourage dense canopy growth and provide moist conditions through increased relative humidity, which is favorable to sporulation. Growers should determine the optimal levels of irrigation at each vineyard site that will result in canopy development that produces desirable yields without excessive shoot or lateral growth.

Figure 13.10 Raisining caused late in the season from Botrytis infection on Riesling. Photo: L. J. Bettiga.
Reduce Berry Damage: Reducing feeding wounds caused by birds, the tortricid moths orange tortrix (Argyrotaenia franciscana) and omnivorous leafroller (Platynota stultana), or other berry-feeding insects can significantly reduce Botrytis bunch rots in coastal areas and diseases caused by other secondary fungal and bacterial pathogens that are more common in the warmer interior valleys. Controlling powdery mildew also reduces berry scarring that can result in cracking during later growth stages, leading to increased levels of bunch rot. Careful monitoring and management of these injuries caused by insects and disease are key components in reducing bunch rot infections. Also, avoid any physical damage to the berries during canopy and cluster management operations.
Chemicals: Lime sulfur has long been used for dormant applications on grapevines. Research has shown that applying lime sulfur at 10 gal/ac (93.6 l/ha) in a high volume of water (at least 100 gal/ac, or 936 l/ha) during the dormant or delayed-dormant state reduces overwintering sclerotia of Botrytis cinerea by 70 to 75%. Lime sulfur kills the sclerotia and significantly reduces inoculum, generally making it a good cleanup product.

Figure 13.11 Lesions and sclerotia on a cane from Botrytis infection. Photo: L. J. Bettiga.
Several fungicide classes are available for in-season control of Botrytis bunch rot. Alternating fungicides that have different modes of action is essential to prevent pathogen populations from developing resistance. Fungicides with a single-site mode of action are more prone to selecting resistant fungus biotypes. Resistance of B. cinerea to benzimidazole (benomyl) fungicide has been observed in California vineyards. Refer to the UC IPM Grape Pest Management Guidelines, www.ipm.ucdavis.edu/ PMG/, for the most current information on registered fungicides for Botrytis bunch rot control.
Spray programs may consist of sprays at bloom or multiple applications at bloom, cluster preclose, veraison, and preharvest. The frequency should depend on the history of Botrytis bunch rot in a particular vineyard, cultivar susceptibility, and the seasonal variation in environmental conditions that are conducive to infection.
Studies to determine the optimal timing of a single application have yielded variable results. A single application at bloom has rarely proved to be significantly more effective than a single application at any other time during the season, the exception being at vineyard sites where there was rain during bloom. At these sites, bloomtime fungicide applications were slightly more effective than at other times. Data collected over many years have shown a direct relationship between the number of fungicide applications and the level of Botrytis bunch rot control when conditions during bloom and after veraison are conducive for infection.

Figure 13.12 Botrytis bunch rot disease cycle.

Figure 13.13 Overwintering Botrytis on mummies (A); spore production on mummies in early spring (B). Photos: J. K. Clark.
Timing sprays according to the plant’s growth stage is usually not effective; a better method is to apply sprays only when environmental conditions conducive to the growth of the fungus have been forecasted. Fungicide applications before rain are more effective in reducing Botrytis infections than those applied after rain. Information on the amount of berry infection resulting from the interaction of temperature and hours of free moisture has been incorporated into commercial software and weather stations, and these can be used to assist in timing applications of fungicides against Botrytis. Real-time or forecasted data can be used to run the Botrytis infection risk model. Generally, the fungicides currently available do not have very long “kick back” activity and must be applied prior to or as soon as possible after a mediumor high-risk event is forecasted.
To obtain optimal coverage with pesticides, spray adjuvants (most commonly surfactants) are often added to the tank mix. The purpose of these surfactants is to reduce the surface tension of the water droplets, allowing better dispersion over the plant’s surface. The nonwettability of a plant part is due mainly to the layer of wax on its surface; some adjuvants actually alter the structure of plant waxes. Although specific situations may call for using spray adjuvants, one must consider that most chemi cals already contain them in their formulation, and that adding more adjuvants can be justified only by data that positively demonstrate a significant increase in efficacy against Botrytis in grape.

Figure 13.14 Strands of fungal tissue develop on berry surfaces. Photo: J. K. Clark.
Plant Resistance: Several factors are involved in a grape berry’s resistance to infection by B. cinerea. Many red cultivars contain compounds that inhibit the fungus. Also, the berry skin provides a mechanical barrier to infection, as does the epicuticular wax on the surface of the berry. Wax formation is inhibited when berries grow in contact with other berries or when the environment is shady and relative humidity is high. These contact areas have been shown to be more susceptible to infection by B. cinerea. In general, tight, thin-skinned white grape berries are the most susceptible to Botrytis bunch rot. However, the thin-skinned red cultivar Zinfandel is also highly susceptible. Although it is unlikely that grape cultivars will ever be developed solely for resistance to Botrytis, a wide range of resistance is already available. See table 13.1 on the relative susceptibility of major grape cultivars in the western United States.
__________________________________________________________________________________
Table 13.1. Relative susceptibility to bunch rot of major grape cultivars in the western United States
| Very susceptible: Carignane Chardonnay Chenin blanc Melon Petite Sirah Ruby Seedless White Riesling Zinfandel |
Susceptible: Barbera Calmeria Flame Seedless Grenache Pinot noir Sauvignon blanc Redglobe Ribier |
Moderately resistant: Autumn Royal Colombard Crimson Seedless Emperor Semillon Sylvaner Thompson Seedless |
Highly resistant: Cabernet Sauvignon Merlot Muscat of Alexandria Rubired Ruby Cabernet |
__________________________________________________________________________________

Figure 13.15 Warm, dry weather at harvest can desiccate Botrytis-infected berries. Photo: L. J. Bettiga.
Research has shown that cluster architecture or tightness may be more important in affecting how much disease develops than the susceptibility of individual berries. Cabernet Sauvignon berries are as susceptible to infection as Zinfandel berries; it is the difference in cluster tightness that influences the microclimate within a Zinfandel cluster that increases Botrytis infection susceptibility. Compact clusters often retain more Botrytis-infected senescent flower parts, and when wetted, the clusters dry more slowly, resulting in a more favorable microclimate for disease infection and spread. Excessively compact clusters also have more physical damage or cracking as berries expand against each other. Clonal evaluation research has shown that selections of Chardonnay and Pinot noir with looser clusters generally have lower levels of bunch rot.
References
- Barbe, G. D., and W. B. Hewitt. 1965. The principal fungus in the summer bunch rot of grapes. Phytopathology 55:815–816.
- Bettiga, L. J., W. D. Gubler, J. J. Marois, and A. M. Bledsoe. 1989. Integrated control of Botrytis bunch rot of grape. California Agriculture 43(2): 9–11.
- Broome, J. C., J. T. English, J. J. Marois, B. A. Latorre, and J. C. Aviles. 1995. Development of an infection model for Botrytis bunch rot of grapes based on wetness duration and temperature. Phytopathology 85:97–102.
- Coley-Smith, J. R., K. Verhoeff, and W. R. Jarvis, eds. 1980. The biology of Botrytis. New York: Academic Press.
- Duncan, R. A., J. J. Stapleton, and G. M. Leavitt. 1995. Population dynamics of epiphytic mycoflora and occurrence of bunch rots of wine grapes as influenced by leaf removal. Plant Pathology 44:956–965.
- English, J. T., C. S. Thomas, J. J. Marois, and W. D. Gubler. 1989. Microclimates of grapevine canopies associated with leaf removal and control of Botrytis bunch rot. Phytopathology 79:395–401.
- Gubler, W. D., J. J. Marois, A. M.
- Bledsoe, and L. J. Bettiga. 1987. Control of Botrytis bunch rot of grape with canopy management. Plant Disease 71:599–601.
- Hewitt, W. B. 1974. Rots and bunch rots of grapes. California Agricultural Experiment Station Bulletin 868.
- Marois, J. J., A. M. Bledsoe, R. M. Bostock, and W. D. Gubler. 1987. Effect of spray adjuvants on the development of Botrytis cinerea on grape berries. Phytopathology 77:1148–1152.
- McClellan, W. D., and W. B. Hewitt. 1973. Time of infection and latency of Botrytis cinerea Pers. in Vitis vinifera L. Phytopathology 63:1151–1157.
- Thomas, C. S., J. J. Marois, and J. T. English. 1988. The effects of wind speed, temperature, and relative humidity on development of aerial mycelium and conidia of Botrytis cinerea on grape. Phytopathology 77:260–265.
- Vail, M. E., and J. J. Marois 1991. Grape cluster architecture and the susceptibility of berries to Botrytis cinerea. Phytopathology 81(2): 188–191.
- Vail, M. E., J. A. Wolpert, W. D. Gubler, and M. R. Rademacher. 1998. Effect of cluster tightness on Botrytis bunch rot in six Chardonnay clones. Plant Disease 82(1): 107–109.
